Simulating Ras: 30 years of progress

Chris Neale is a Staff Scientist at the Los Alamos National Laboratory in New Mexico. His research focuses on computational drug discovery and membrane protein biophysics.
The first crystal structures of Ras arrived more than thirty years ago.1,2 Within only a few years, its structure and dynamics were being simulated on computers.3,4 As with many things in science, progress was rapid. Whereas these early simulations probed solution systems on sub-nanosecond time scales, recent ensembles of simulations of Ras protein(s) on mixed lipid bilayers attained aggregate time scales exceeding a millisecond using all-atom models,5 and exceeding 200 milliseconds using coarse-grained models in which each protein residue is represented by 2-5 beads.6 Together, computer simulations and associated theoretical modeling approaches have made important contributions to our current understanding of Ras’s disposition with respect to the membrane surface and the nature of Ras’s interactions with its effector kinase Raf. Both of these advances relate to druggable interactions.
Membrane orientation of Ras
The three-dimensional orientation adopted by a rotatable peripheral membrane protein has the potential to impact signaling-related protein-protein interactions (Fig. 1). For Ras, relevant protein binding sites include those for regulators, effectors, and, possibly, interfaces involved in dimerization and/or multimerization.7 Unlike binding interactions between freely-diffusing cytosolic proteins, for which orientation is unlikely to be relevant (Fig. 1A), peripheral membrane proteins such as Ras may adopt orientations that expose (Fig. 1B) or hide (Fig. 1C) particular binding sites. The latter condition, membrane occlusion, may arise when binding site(s) on Ras are in direct membrane contact, or when they are sufficiently close to the membrane that the space required by a binding partner is unavailable.
If the reorientation of Ras occurs in a manner that is independent from environmental influence, it is reasonable to treat the signaling interference from membrane occlusion as a non-specific crowding effect.8 However, if Ras’s membrane orientation is modulated by local lipid concentration or other environmental conditions, it becomes useful to consider the membrane as a competitive inhibitor, whose on- and off-rates are controlled by the (druggable) dynamics of G domain reorientation.
One additional means of regulation, currently lacking any direct evidence for Ras, involves the potential for membrane-based induction of protein orientations that are pre-arranged to facilitate multimerization or other types of protein-protein interactions between two or more peripheral membrane proteins (Fig. 1D). In many ways, such pre-organization is the converse of membrane-based occlusion, and can be thought of in terms of membrane action as a positive allosteric modulator.
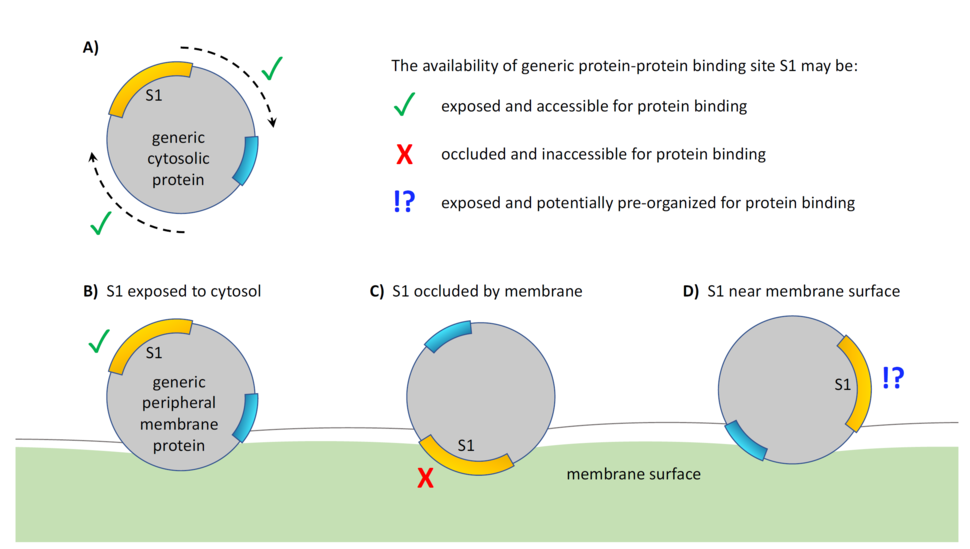
Figure 1: Protein-protein interactions of rotatable peripheral membrane proteins can be affected by their orientation with respect to the membrane surface. (A) The absolute orientation of a monomeric, uncomplexed cytosolic protein does not generally affect its ability to interact with other such proteins. Conversely, the orientation of a peripheral membrane protein does have the potential to affect this type of interaction based on a protein binding interface that is either (B) exposed, (C) occluded, or (D) in a subset of the exposed state that is pre-arranged to facilitate binding interactions.
A number of computational studies have characterized statistical ensembles of the membrane orientation adopted by the G domain of H-Ras,9,10 N-Ras,11 K-Ras4A,12 and K-Ras4B.5,6,13-20 Together with experiments, these computational analyses of Ras’s membrane orientation, pioneered by Gorfe and McCammon,9 have revealed apparent orientational dependencies on isoform,21 oncogenic mutation,17 nucleotide loading,9,10,17,18,22 and local lipid composition.5,6,12,20 Here, experimental evidence is consistent with such correlations, but the nature of experimental biochemical inquiry frequently leads to some ambiguities in structural interpretation. Conversely, simulation models provide exceptional resolution, but are subject to substantial uncertainties due to relatively short time scales and, more often than not, insufficient simulation repetition. As computational resources continue to mature, large ensembles of simulations are becoming more common5,6,20 and offer the potential to overcome these statistical limitations. Indeed, the combination of data from multiple sources has led to a convergent picture of K-Ras4B membrane interactions in which anionic lipids lead the G domain to contact the membrane predominantly via helices 4 and 5 (a4/5 state), or helices 3 and 4 (a3/4 state), or to tilt toward b sheets 1-3 (b1/2/3 state).5
Arguably the most important outcome from the last decade of computational studies on the membrane orientation of Ras is the development of a small molecule that reduces the binding of nanodisk-tethered K-Ras to the Ras binding domain (RBD) of B-Raf in a concentration dependent manner. This compound, presented by Fang and Ikura,23 appears to act by bridging the G domain to the membrane in such a way so as to stabilize orientations in which Ras’s effector binding site is occluded by the membrane. As such, it represents an exciting new avenue in the development of Ras inhibitors based on membrane occlusion.
Quaternary arrangement in Ras-Raf complexes
Structures of the Raf-1 RBD24-26 and cysteine rich domain (CRD)27 have been available for more than 20 years, as has a signaling-relevant structure of the Raf-1 RBD in complex with a Ras homolog, Rap1A.28 The first structural characterization of Ras bound to the Raf-1 RBD is more recent,29 and the last year has delivered two crystal structures of the complex between the tandem RBD/CRD domains of Raf-1 and K-Ras30 or H-Ras.31 Both of these structures reveal a similar quaternary organization in which the CRD’s alpha helix packs against Ras. Using Ras orientations commonly observed in all-atom molecular dynamics simulations, Tran and Simanshu30 showed that only the a4/5 orientational state of Ras puts the CRD’s hydrophobic/cationic membrane-active loops32,33 in contact with the membrane in the context of these crystallographic structures (Fig. 2A). Although the a3/4 orientational state of Ras appears to be incompatible with membrane interactions of either the RBD or CRD (Fig. 2B), NMR-driven models33 suggest that the b1/2/3 orientational state of Ras is also compatible with membrane insertion of the CRD’s loops if the CRD is rotated by approximately 180° about the RBD-CRD long axis with respect to available solution-state crystal structures,30,33 despite the moderate inconsistency of such interactions with NMR data32,33 using models based on available crystal structures (Fig. 2C).
Although the crystallographic Ras-CRD interface has an area similar to that of the Ras-RBD interface and is stabilized by a number of hydrogen bonds,30 simulations of K-Ras4B bound to the B-Raf RBD/CRD, conducted prior to the emergence of crystal structures or NMR-driven models of this complex, revealed the potential for multiple distinct modes of Ras-CRD interaction.32 Moreover, these simulations showed concerted membrane interaction of Ras, the RBD, and the CRD.32 Clearly, membrane interactions of the CRD and their relation to Ras orientation remains an area of active interest, as does the evaluation of possible higher-order complexes.34
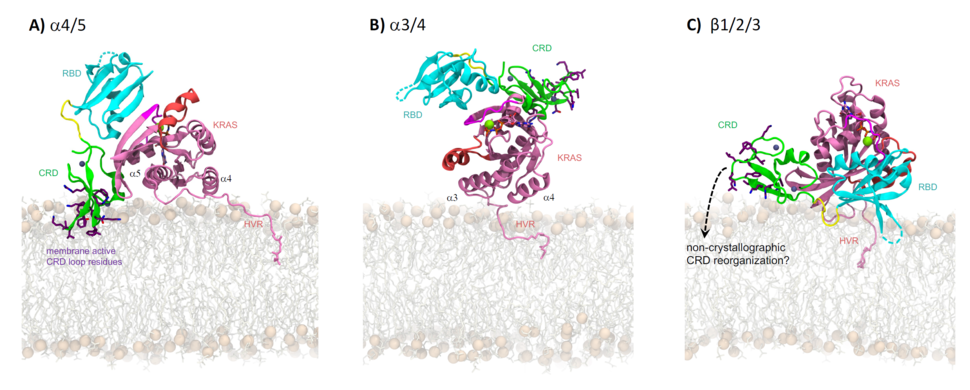
Figure 2: Orientations of Ras from computer simulations and their impact on the membrane interactions of the crystallographic pose of the Ras-RBD/CRD complex. Ras is colored pink with switch-I and switch-II regions highlighted in magenta and salmon, respectively. RBD, CRD, and the linker present between them are colored cyan, green and yellow, respectively. Side chains of CRD residues R143KTFLKLAF151 and K157FLLNGFR164 are sticks with purple carbon atoms. Mg2+ (green) and Zn2+ (gray) are shown as spheres. Lipids are sticks with spheres for head group phosphorus atoms. Figure adapted from Tran et al.30